論文紹介LUCUBRATIONS
Differential vagal effects on antegrade vs.retrograde atrioventricular conduction
Cardiovascular Division, The Lankenau Medieal Research Center, and Department of Medicine, Jefferson Medical Collcge of Thomas Jefferson University, Philadelphia, Pennsylvania 19151; and Bulgarian Academy of Sciences, Sofia, Bulgaria
TAKAO MITSUOKA, TODOR MAZGALEV, LEONARD S. DREIFUS, AND ERIC L. MICHELSON. Differential vagal effects on antegrade vs. retrograde atrioventricular conduction. Am. J.Physiol. 253 (Heart Circ. Physiol. 22): H1059 - H1068, 1987 -The infuluence of postganglionic vagal stimulation (PGVS) on antegrade and retrograde atrioventricular nodal conduction was studied in 17 isolated rabbit heart tissue preparations by pacing at the crista terminalis or His bundle, respectively. The effect of short bursts of PGVS on prolongation of atrioventricular conduction was phase dependent with respect to the cardiac cycle. This phasic dependency was more pronounced during antegrade atrioventricular conduction. Although the control retrograde atrioventricular conduction time was longer than the antegrade (P < 0.05) at or near the time in the cycle during which vagal stimulation caused maximal prolongation of conduction time (optimal phase), PGVS-induced maximal prolongation of the antegrade atrioventricular conduction time was significantly greater than that of the retrograde (P < 0.02). Moreover, when PGVS was introduced at a fixed phase in the cycle, but with increasing amplitude, antegrade atrioventricular conduction time was progressively prolonged, and block was observed first in the antegrade direction, whereas retrograde atrioventricular conduction continued. Microelectrode recordings during these experiments showed consistently that PGVS-induced hyperpolarization in the N region of the atrioventricular node was greater during antegrade atrioventricular conduction. This suggests that vagal effects depended not only on the intensity and phase of stimulation, but also on electrotonic influences which apparently are different during antegrade and retrograde conduction.
postganglionic vagal stimulation; rabbit; N region
ALTHOUGH THE DEPRESSIVE EFFECTS of vagal stimulation on atrioventricular nodal conduction have been intensively studied, little is known about the influence of the direction of impulse propagation in the atrioventricular node (AVN) on the effect of the vagus.
Previous studies6) 8) done in isolated tissue preparations showed that acetylcholine exerts different effects on antegrade and retrograde AVN activation. Recently, several clinical studies have shown that atropine can restore unidirectionally blocked retrograde AVN conduction, and these investigators concluded that high vagal tone might be responsible for transient (or intermittent) retrograde conduction block 3) 13) 21). These studies also suggested that vagal effects on the AVN could differ depending on the direction of impulse propagation.
This study was designed to characterize the effects of postganglionic vagal stimulation (PGVS) applied in the AVN on 1) changes in antegrade vs. retrograde atrioventricular nodal conduction time, and 2) the corresponding changes in cellular action potentials in the different regions of the AVN.
METHODS
The preparations used and the methods of electrical stimulation and recording were similar to those previously described 14) 16). All experiments conformed to the National Institutes of Health guide for Care and Use of Laboratory Animals and the Lankenau Animal Care Policy. In brief, atrial-atrioventricular nodal preparations from 17 New Zealand White rabbits with body weights of ~2.5 kg were used. As detailed below, extracellular recordings were done in all 17 preparations and, in addition, microelectrode studies were done in seven of these preparations. The preparations consisted of the sinoatrial nodal region, crista terminalis (CrT), interatrial septum (IAS), AVN, and the proximal portion of the bundle of His (H). The preparations were mounted endocardium-up on a 5-mm thick silicon disc and placed in a tissue bath for superfusion at a constant flow rate of 20 ml/min with Tyrode's solution at 35.0°C. The Tyrode's solution was saturated with a mixture of 95% O2-5% CO2 and had the following composition (concentrations in mM): NaCl 128.2, KCl 4.7, CaCl2 1.3, MgCl2 1.05, NaHCO2 20.2, NaH2PO4 0.7, and dextrose 2 g/l; pH was 7.25-7.40. Propranolol hydrochloride (1.5 mg/l) was used as a β-adrenergic blocking agent.
Bipolar surface platinum electrodes were used for recording from the CrT near the posterior input of the AVN, from the IAS near the anterior input of the AVN, and from the His bundle.
To study antegrade and retrograde AVN activation, the preparations were driven by electrical stimulation with a constant cycle length (400-500 ms) and an amplitude of twice diastolic threshold, delivered through bipolar electrodes incorporated with recording surface electrodes at the CrT or His bundle regions. Antegrade and retrograde AVN conduction times were measured as intervals between the CrT and His electrograms, or His and CrT electrograms, respectively. These times were determined electronically using the first rapid reflection exceeding a predetermined threshold value for each electrogram. The time resolution was 1 ms.
Standard glass microelectrodes (Kwik-Fil, World Precise Instruments) with a resistance of 20-40 M were used to record (S7100A Modular Microprobe System, World Precise Instruments) action potentials from AVN cells. Recordings were obtained from the same impaled cell during antegrade and retrograde activation for comparison. Single short bursts (duration 50-100 ms) of impulses with subthreshold intensity for the atria, a pulse width of 1 ms and interimpulse intervals of 6 ms, were delivered through a bipolar platinum electrode to produce PGVS of the AVN 29). The position of the PGVS electrode (except when specifically mentioned) was in or near the N region of the AVN where the maximal effect was observed during both antegrade and retrograde conduction. However, qualitatively the phenomenon observed did not depend on the precise position of the PGVS electrode. The amplitude of the impulses in the bursts was empirically determined so that the dromotropic effects were ~30-50% of the maximum. The usual range of these amplitudes was 70-125 μA. To observe AVN block, the amplitude of PGVS was increased to 150-500 μA. The burst generator 16) and a programmable stimulator (Bloom Associates) were synchronized with the atrial (CrT) or His electrogram. The vagal bursts were introduced with a programmed delay to scan the cycle length in incremental 50-ms steps. The technical devices used in the experiments were previously described in detail 14) 16).
The arrangement of the traces shown in the figures is (from top to bottom): electrograms from the CrT and IAS, action potential(s) recorded from AVN cell(s), and the His electrogram. The timing of PGVS is marked by a horizontal bar above the H tracing. The position of PGVS in the cycle was determined by the absolute phase (APh): the time interval between the preceding electrogram (atrial or ventricular for antegrade and retrograde stimulation, respectively) and the vagal burst 16). If not mentioned specifically, AV nodal conduction time refers to the first beat after PGVS.
Statical Analysis
Wilcoxon's signed-rank test was used to compare antegrade and retrograde AVN conduction times before and after PGVS in composite data. Results presented below from selected experiments, and specifically the microelectrode data, were typical of all preparations unless otherwise noted.
RESULTS
Effect of PGVS on AVN Conduction Time During Antegrade and Retrograde Activation
The effects of PGVS on AVN conduction time in the various regions of the AV node are illustrated in Fig. 1. In this experiment, the preparation was driven at a cycle length of 400 ms. The electrode for PGVS was sequentially positioned in different regions of the node (Fig. 1, circles). PGVS was introduced with the following parameters: burst duration 50 ms, APh 100 ms, and amplitude 300 μA. The circled numbers in Fig. 1 represent the changes (in milliseconds) in the antegrade (top numbers) and retrograde conduction times (bottom numbers) when the stimulating electrode was positioned at each of these respective points in the node. These changes were calculated as the difference between the conduction time of the first beat after PGVS and the basic conduction time before PGVS. As shown, the maximal prolongation of the conduction time occurred with positioning of the stimulating electrode in the N and NH regions of the node during both antegrade and retrograde activation. PGVS introduced in either the AN or His bundle regions produced little or no change in conduction time in either the antegrade or retrograde direction. A high amplitude of PGVS was used for illustrative purposes in Fig. 1 to produce AVN block and identify the effects of vagal stimulation in the various regions of the AVN. However, lower amplitudes, preserving 1:1 AVN conduction, were utilized to demonstrate the different phasic effects of PGVS during antegrade vs. retrograde conduction.
FIG. 1.
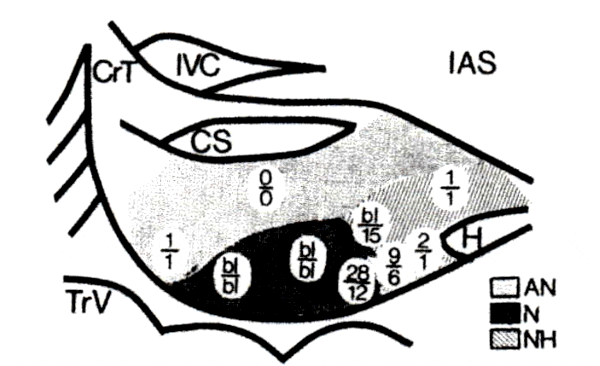 Comparison of postganglionic vagal stimulation (PGVS)- induced conduction delay measured with PGVS-stimulating electrode at different locations within the atrioventricular node (AVN), CrT, crista terminalis; IAS, interatrial septum; IVC, inferior vena cava; CS, coronary sinus; TrV, tricuspid valve; H, His bundle. Sh&d areas represent approximately AN region, N region, and NH region. Numbers in circles represent AVN conduction time, antegrade above the lines and retrograde below, respectively, after introducing same PGVS through an electrode located in each respective circled area. bl, block of AVN conduction. Note that with both antegrade and retrograde activation the major delay in conduction time was observed in the N region. See text for details.
Comparison of postganglionic vagal stimulation (PGVS)- induced conduction delay measured with PGVS-stimulating electrode at different locations within the atrioventricular node (AVN), CrT, crista terminalis; IAS, interatrial septum; IVC, inferior vena cava; CS, coronary sinus; TrV, tricuspid valve; H, His bundle. Sh&d areas represent approximately AN region, N region, and NH region. Numbers in circles represent AVN conduction time, antegrade above the lines and retrograde below, respectively, after introducing same PGVS through an electrode located in each respective circled area. bl, block of AVN conduction. Note that with both antegrade and retrograde activation the major delay in conduction time was observed in the N region. See text for details.
A typical PGVS-AVN conduction time-phasic response curve showing the relationship between the conduction time and the absolute phase of PGVS is illustrated in Fig. 2. This preparation was driven at a cycle length of 400 ms. The control retrograde AVN conduction time (116 ms) was longer than the antegrade AVN conduction time (90 ms). The effect of PGVS (amplitude 75 μA, duration 50 ms) on the prolongation of AVN conduction time was phase dependent with a maximum occurring at 100 ms APh. This phasic dependency existed for both antegrade (open circles) and retrograde (filled circles) AVN conduction, but was more pronounced for antegrade AVN conduction. Consequently, during most phases of PGVS, antegrade AVN conduction time became longer than retrograde conduction time. Notably, even when PGVS amplitude was increased to 300 μA (not shown), retrograde conduction was additionally prolonged only slightly, whereas block was observed during antegrade conduction.
FIG. 2.
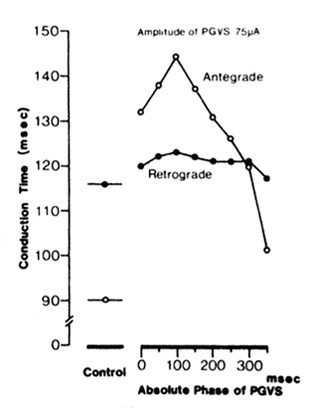 A typical phasic response curve of atrioventricular nodal (AVN) conduction time, i.e., relationship between conduction time and absolute phase of postganglionic vagal stimulation (PGVS) (duration 50 ms, amplitude 75 μA). Open circles, antegrade AVN conduction; filled circles, retrograde AVN conduction. Preparation was driven at a cycle length of 400 ms. Note that during control, retrograde AVN conduction time was longer than antegrade AvN conduction time, whereas during most phases of PGVS, antegrade AVN conduction time was longer than retrograde AVN conduction time.
A typical phasic response curve of atrioventricular nodal (AVN) conduction time, i.e., relationship between conduction time and absolute phase of postganglionic vagal stimulation (PGVS) (duration 50 ms, amplitude 75 μA). Open circles, antegrade AVN conduction; filled circles, retrograde AVN conduction. Preparation was driven at a cycle length of 400 ms. Note that during control, retrograde AVN conduction time was longer than antegrade AvN conduction time, whereas during most phases of PGVS, antegrade AVN conduction time was longer than retrograde AVN conduction time.
FIG. 3.
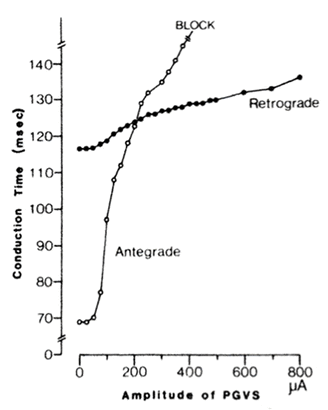 Effect of increasing amplitude of postganglionic vagal stimulation (PGVS; duration 50 ms) with a fixed absolute phase (Aph; 150 ms) on antegrade and retrograde atrioventricular nodal (AvN) conduction times in a typical preparation, which was driven at a cycle length of 450 ms. Note that increasing amplitude of PGVS resulted in a greater prolongation of antegrade (open circles) than retrograde AVN conduction (filled circles) time. At amplitudes of PGVS > 200 μA, antegrade conduction time was longer than retrograde. AVN block was also observed only during antegrade conduction within this range of stimulation amplitudes.
Effect of increasing amplitude of postganglionic vagal stimulation (PGVS; duration 50 ms) with a fixed absolute phase (Aph; 150 ms) on antegrade and retrograde atrioventricular nodal (AvN) conduction times in a typical preparation, which was driven at a cycle length of 450 ms. Note that increasing amplitude of PGVS resulted in a greater prolongation of antegrade (open circles) than retrograde AVN conduction (filled circles) time. At amplitudes of PGVS > 200 μA, antegrade conduction time was longer than retrograde. AVN block was also observed only during antegrade conduction within this range of stimulation amplitudes.
When PGVS was introduced at a fixed APh in the cycle, but with increasing amplitude (Fig. 3), the antegrade AVN conduction time (open circles) was progressively prolonged, and block was observed first in the antegrade direction, whereas retrograde AVN conduction (filled circles) continued despite a high amplitude of PGVS. At amplitudes of PGVS > 200 μA, antegrade AVN conduction time was longer than retrogade AVN conduction time.
A summary of the data from the seven preparations in which only extracellular recordings were done is shown in Fig. 4 to demonstrate the effect of PGVS on antegrade and retrograde AVN conduction. The preparations were driven at a basic cycle length of 400-450 ms. As previously described, the cycle length was scanned by PGVS bursts, and the maximally prolonged AVN conduction time. in each preparation is represented in Fig. 4. The maximally prolonged antegrade and retrograde conduction times were observed at similar absolute phases of PGVS in a given preparation. Note that in the control studies (Fig. 4, left) the antegrade conduction time in six and similar in one preparation. This tendency was reversed with a “low” intensity PGVS (amplitude 70-125 μA) in three of the preparations (Fig. 4, middle). Moreover, it can be concluded from the presented data that the relative changes in AVN conduction vs. control in this instance were greater during antegrade conduction in each of the seven preparations, “High" intensity PGVS (amplitude 125 -500 μA, Fig. 4, right) produced block in all preparations during antegrade but not retrograde conduction. Thus the increase of AVN conduction time was significantly greater during antegrade the retrograde conduction (P < 0.02). Consequently, although the control retrograde AVN conduction time was significantly (P < 0.05) longer than the antegrade AVN conduction time (Fig. 4, left), increase vagal tone produced a longer antegrade AVN conduction time that retrograde AVN conduction time (Fig. 4, middle and right).
FIG. 4.
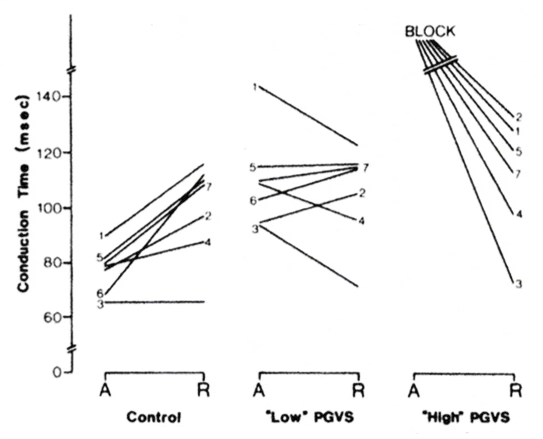 Summary of data from 7 preparations (numbers 1-7) demonstrating effect of postganglionic vagal stimulation (PGVS; duration 50 ms) on atrioventricular nodal (AVN) conduction time. Preparations were driven at a cycle length of 400-450 ms. Only maximal effects of PGVS, i.e., those observed at optimal phase, are shown. Absolute changes in AVN conduction times (A, antegrade; and R, retrograde) resulting from PGVS with "low" (middle panel) or “high" (right panel) amplitude. Amplitude levels were determined empirically in each preparation so that “high” was associated with AVN block, whereas “low” resulted in conduction delay but not block. Note that absolute and relative changes in conduction times as compared with control (left) were accentuated during antegrade conduction. See text for details.
Summary of data from 7 preparations (numbers 1-7) demonstrating effect of postganglionic vagal stimulation (PGVS; duration 50 ms) on atrioventricular nodal (AVN) conduction time. Preparations were driven at a cycle length of 400-450 ms. Only maximal effects of PGVS, i.e., those observed at optimal phase, are shown. Absolute changes in AVN conduction times (A, antegrade; and R, retrograde) resulting from PGVS with "low" (middle panel) or “high" (right panel) amplitude. Amplitude levels were determined empirically in each preparation so that “high” was associated with AVN block, whereas “low” resulted in conduction delay but not block. Note that absolute and relative changes in conduction times as compared with control (left) were accentuated during antegrade conduction. See text for details.
Effect of PGVS on AVN potentials Cell Action Potentials
PGVS-induced hyperpolarization during antegrade vs. retrograde conduction. The phasic effect of PGVS corresponded to the degree of AVN hyperpolarization at the moment that atrial or His bundle activation arrived at the AVN 14). This is illustrated in Fig. 5. The action potentials were recorded from a cell in the N region of the AVN. PGVS (duration 50 ms, amplitude 150 μA) was introduced at phases of 50, 150, 250, and 350 ms (as measured from the last CrT or H electrogram in antegrade or retrograde mode, respectively). PGVS with 150 ms APh was associated with antegrade AVN block (Fig. 5A, second panel from the left, straight arrow), whereasPGVS introduced either earlier (50ms) or later (250 and 350 ms) was associated with prolongation of the conduction time. It is also clear that the phasic effects of PGVS on the antegrade conduction time reflected change,s in the membrane potential. At an APh of 50 ms, PGVS caused hyperpolarization of the membrane; the marked depression of the action potential amplitude suggested only local response (Fig. 5A, first panel from the left, filled curved arrow). However, despite the local block, conduction continued toward the His bundle (filled straight arrow). Hence, conduction occurred presumably using other fibers which were less influenced by the PGVS. The second hump of the membrane potential was recorded after the inscription of the His bundle electrogram, suggesting local reentry in this region of the AV node. When PGVS was moved 100 ms later, namely at an APh of 150 ms (Fig. 5A, second panel from the left), PGVS caused further hyperpolarization of the membrane, a local response without the second hump (open curved arrow), and an absence of H potential (straight thin arrow) associated with AVN block. When PGVS was moved even later (APh = 250 or 350 ms), the influence on antegrade conduction through the AVN was less; PGVS caused less hyperpolarization, a less negative take-off, and subsequently less depression of the amplitude and the first time derivative (dV/dt) of the action potential. Note that PGVS during later phases had a gradually increasing effect on the conduction of the subsequent beat (Fig. 5A, asterisks). Figure 5B shows the changes in the action potentials after PGVS during retrograde AVN conduction. In contrast to antegrade conduction, PGVS with APh of 50 or 150 ms (Fig. 5B, first and second panels from the left) caused less hyperpolarization and less depression of action potential duration, amplitude, and dV/dt in comparison with the antegrade mode. When PGVS was moved to a later phase (250 or 350 ms), the changes observed in this impaled N cell were almost negligible. This corresponded to the observation that overall there was less phasic dependency of the conduction time during retrograde than antegrade activation (see also Fig. 2A).
FIG. 5.
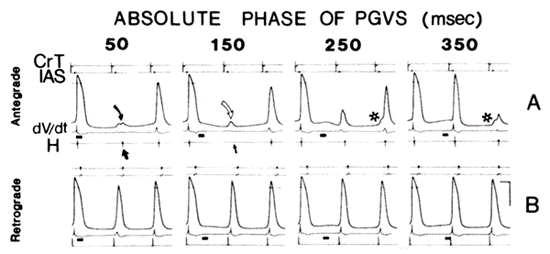 Phasic effects of postganglionic vagal stimulation [PGVS; horizontal short bar above His bundle (H) tracing, duration 50 ms, amplitude 150 μA] during antegrade (A) and retrograde (B) conduction. Preparation was driven at a cycle length of 400 ms. In all panels from top to bottom: crista terminalis (CrT) electrogram, lower interatrial septum (IAS) electrogram, action potentials recorded from atrioventricular node (AVN) cell, dV/dt which represents first time derivative of the action potential, and His bundle electrogram (H). Thin broken line here and in later action potential recordings represents zero membrane potential. Note the greater effect of similar PGVS bursts on action potential characteristics during antegrade than retrograde conduction. *Beats subsequent to introduction of PGVS. See text for details about other symbols. Calibration: 100 ms (horizontal), 50 mV (vertical, for action potentials) and 30 V/s (vertical, for dV/dt).
Phasic effects of postganglionic vagal stimulation [PGVS; horizontal short bar above His bundle (H) tracing, duration 50 ms, amplitude 150 μA] during antegrade (A) and retrograde (B) conduction. Preparation was driven at a cycle length of 400 ms. In all panels from top to bottom: crista terminalis (CrT) electrogram, lower interatrial septum (IAS) electrogram, action potentials recorded from atrioventricular node (AVN) cell, dV/dt which represents first time derivative of the action potential, and His bundle electrogram (H). Thin broken line here and in later action potential recordings represents zero membrane potential. Note the greater effect of similar PGVS bursts on action potential characteristics during antegrade than retrograde conduction. *Beats subsequent to introduction of PGVS. See text for details about other symbols. Calibration: 100 ms (horizontal), 50 mV (vertical, for action potentials) and 30 V/s (vertical, for dV/dt).
PGVS effects in different regions of AVN during antegrade vs. retrograde conduction. The N region of the AVN was apparently the major focus of vagal influence during both antegrade and retrograde conduction. Figure 6 illustrates the effects of PGVS on AVN cells from three regions of the AVN: AN, N, and NH. In Fig. 6, A-D, action potentials were recorded from the AN, distal AN (near N), N, and NH regions, respectively, as shown in the inset. The results in Fig. 6, A, B, and D, were obtained from the same preparation driven at a basic cycle length of 450 ms, whereas Fig. 6C was obtained from a different preparation with a basic cycle length of 400 ms. Each panel represents antegrade (top records) and retrograde (bottom records) conduction, and the records of action potentials were obtained from the same cells during antegrade and retrograde conduction. Several increasing amplitudes of PGVS were utilized to demonstrate the development of antegrade and/or retrograde AVN block. A PGVS burst was introduced with a duration of 50 ms, APh at 150 ms in Fig. 6 A, B, and D, and with a duration of 50 ms, APh at 100 ms in Fig. 6C. In all cases shown the absolute phase was optimal, i.e., PGVS effect was maximal for the given amplitude in both the antegrade and retrograde directions.
At the top of Fig. 6A, antegrade stimulation depolarized the AN cell after PGVS, but distal to this cell AVN block occurred (no His bundle activation as shown by a thick straight arrow). Stimulation of the His bundle after PGVS (Fig. 6A, bottom panel) was followed by retrograde block distal to the same impaled cell. A spontaneous sinus discharge (asterisk) was responsible for the antegrade conduction and His bundle electrogram inscription, thus preventing subsequent retrograde activation of the His bundle (the stimulus artifact is indicated by the thin arrow). Note that the amplitude of PGVS (300 μA) was sufficient to produce AVN block distal to this AN cell during both antegrade and retrograde stimulation. In Fig, 6B, top panel, antegrade conduction caused deporariation of the impaled distal AN cell (inset, point B) after PGVS(180 μA) but was blocked distal to this cell (no His bundle activation as shown by the filled, straight arrow). However, the same PGVS could not induce retrograde AVN block (Fig. 6B, bottom middle panel, (althought prolongation of retorgrade AVN conduction and distortion of the action potential (filled curved allow) were observed. After inquiresing the amplitude to 300 μA, (Fig. 6B, bottom right panel). retrograde AVN block occured distal to this cell after PGVS, and only a local respose was observed (open curved arrow). As a result, the spontaneous sinus beat (asterisk) was conducted antegradely followed by His bundle acetiction. In Fig. 6C, top panel, the increasing amplitude of PGVS (50, 75, and 300 μA) was associated with a gradual deterioration of the action potential of the N cell (curved arrows) and subsequently with depressed AVN conduction. After PGVS with an amplitude of 180 μA (not shown) as well as 300 μA (Fig. 6C, top right panel) antegrade conduction was blocked (straight arrow). In contrast, retrograde conduction (Fig. 6C, bottom panels) was less influenced by PGVS. With amplitudes up to 300 μA, the deterioration of the action potential recorded from the same cell was less than n in the integrated mode. only at 300 μA (bottom right panel) did PGVS produce retrograde AVN block, presumably in the vincinty of the impaled cell. This retrograde block permitted the next spontaneous sinus nodal discharge (asterisk) to be conducted antegradely. Finally, as shown in Fig. 6D, top panel, antegrade activation was blocked to after PGVS proximal to this NH cell. which showed a local response (open curved arrow) without His activation (filled straight arrow). However, the same amplitude of PGVS was not sufficient to induce retrograde AVN block (Fig. 6D, bottom middle panel). A higher amplitude of PGVS (300 μA Fig. 6D, bottom right panel) produced retrograde AVN block proximal to this cell. As a result, the spontaneous sinus beat (asterisk) depolarized the cell but His activation was not seen, presumably due to refractoriness of a more distal portion of the AVN.
Antegrade as well as retrograde AVN block cured in the region distal to cells A and B, but proximal to cell D. Namely, the most significant influence of PGVS was observed in cells of the N region (Fig. 6C) of the AVN during antegrade conduction, whereas AN cells (Fig. 6, A and B) were not influenced. During retrograde conduction, the major effect of PGVS was also observed in the cells from the N region (Fig. 6C), whereas distal NH cells (Fig. 6D) and AN cells (Fig. 6, A and B) were influenced little. A higher PGVS amplitude was always required to induce retrograde AVN block in comparison with antegrade AVN block (300 vs. 180μA in this instance).
FIG. 6.
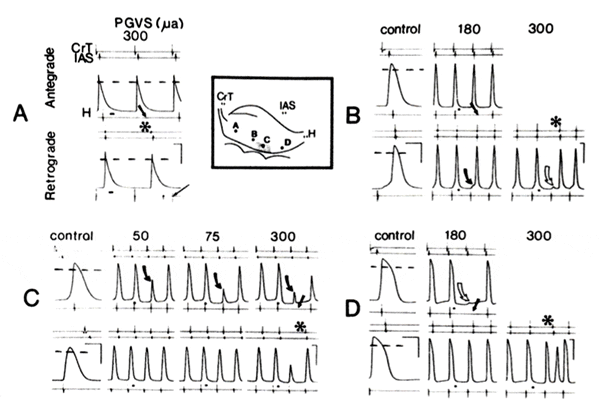 N region of atrioventricular node (AVN; shaded area in the inset) was major focus of vagal influence during both antegrade (top records in each panel) and retrograde (bottom records) conduction. A-D, action potentials were recorded from different AVN cells at marked by corresponding letters in the inset. Control recordings demonstrate changes in shape of action potentials during antegrade vs. retrograde conduction. A, B, and D were obtained from same preparation, whereas C was from an additional preparation in which 3 amplitudes of postganglionic vagal stimulation (PGVS) were used. In each panel several increasing amplitudes of PGVS (Aph 150 ms, duration 50 ms in A, B, and D, and APh 100 ms, duration 50 ms in panel C) were utilized to demonstrate AVN block in both directions. Note that antegrade AVN block occurred in region distal to celsl A and B, but proximal to cell D, whereas retrograde AVN block occurred in region distal to cell D but proximal to cells A and B. AVN block in both directions occured in region of cell C (N region). See text for details about other symbols. Calibration: 100 ms (horizontal) and 50 mV (vertical, for action potentials).
N region of atrioventricular node (AVN; shaded area in the inset) was major focus of vagal influence during both antegrade (top records in each panel) and retrograde (bottom records) conduction. A-D, action potentials were recorded from different AVN cells at marked by corresponding letters in the inset. Control recordings demonstrate changes in shape of action potentials during antegrade vs. retrograde conduction. A, B, and D were obtained from same preparation, whereas C was from an additional preparation in which 3 amplitudes of postganglionic vagal stimulation (PGVS) were used. In each panel several increasing amplitudes of PGVS (Aph 150 ms, duration 50 ms in A, B, and D, and APh 100 ms, duration 50 ms in panel C) were utilized to demonstrate AVN block in both directions. Note that antegrade AVN block occurred in region distal to celsl A and B, but proximal to cell D, whereas retrograde AVN block occurred in region distal to cell D but proximal to cells A and B. AVN block in both directions occured in region of cell C (N region). See text for details about other symbols. Calibration: 100 ms (horizontal) and 50 mV (vertical, for action potentials).
The above detailed findings were consistent in six of seven preparations in which microelectrode studies were performed. Only one preparation showed a dissimilar behavior. In this instance, retrograde AVN block was induced by a PGVS burst, which did not produce antegrade AVN block, as illustrated in Fig. 7. The preparation was driven at a cycle length of 450 ms. The short bar above the H electrogram (Fig. 7) represents PGVS (APh = 250 ms, duration 100 ms, amplitude 50 μA), and the numbers below the H electrogram indicate the conduction time from the CrT to the His bundle and vice versa.
In Fig. 7A, a small degree of hyperpolarization was observed after PGVS in an AN cell and was associated with a moderate prolongation of the antegrade conduction time (from 87 to 96 ms). In contrast, during retrograde conduction the same AN cell showed no hyperpolarization after the same PGVS, but fragmentation of the action potential was observed within several beats associated with a greater prolongation of the retrograde conduction time (from 98 to 133 ms in the first cycle and to 145 ms in the second cycle, Fig. 7B).
In Fig. 7C, an additional action potential recording (the fourth trace from the top) revealed hyperpolarization of an N cell in this experiment. The amplitude of PGVS was increased from 50 to 100 μA, otherwise similar parameters of PGVS and the basic cycle length were used. In Fig. 7C, typical hyperpolarization of the N cell after PGVS was associated with a greater prolongation of the conduction time (from 87 to 112 ms) in comparison with Fig. 7A (from 87 to 96 ms). During retrograde conduction (Fig. 7D), the same N cell showed less hyperpolarization after PGVS. Despite this, a greater prolongation of the conduction time (from 98 to 137 ms in the first cycle) was observed. Moreover, retrograde AVN block occurred in the second cycle after PGVS and was apparently located proximal to the impaled N cell and close to the AN cell. The subsequent spontaneous sinus beat (asterisk) was conducted antegradely. Consequently, retrograde pacing (arrowhead) failed to activate the His bundle due to refractoriness in this area after the antegrade propagation. Thus the observed increase in PGVS effect on retrograde conduction in this caEe was not related to a greater hyperpolarization in the N region. Rather, inhomogeneous conduction through the AN region during retrograde stimulation was induced by PGVS and accounted for the AVN block.
FIG. 7.
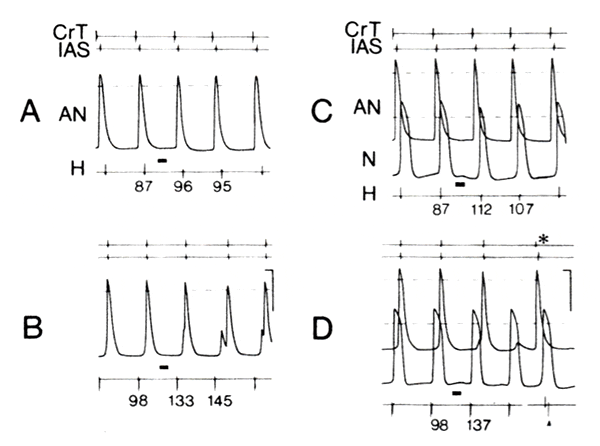 A and B: an atypical preparation in which retrograde AVN conduction was influenced to a greater degree by postganglionic vagal stimulation (PGVS) than antegrade atrioventricular nodal (AVN) conduction. Organization of records was as follows (from top to bottom): crista terminalis (CrT) electrogram, lower interatrial septum (IAS) electrogram, action potentials from AN region, and His bundle electrogram (H). Snort bar above H electrogram represents PGVS (APh 250 ms, duration 100 ms, amplitude 50 μA) and numbers below the H electrogram indicate conduction time (in ms). Preparation was driven at a cycle length of 450 ms. A shows antegrade conduction and I retrograde conduction. Note that fragmentation of action potential was induced by PGVS during retrograde conduction (B) which could account for longer retrograde that antegrade conduction time. C and D: same preparation and organization of records as A and B. In addition, a simultaneous recording of action potential from an N cell was added and amplitude of PGVS was incresed to 100 μA. Note that PGVS-induced hyperpolarization in N cell was greater during antegrade conduction (C). In contrast, the AVN conduction (D), and AVN block was observed distal to the impaled AN cell during the second cycle after PGVS. *Spontaneous sinus beat. See text for details. Calibration: 100 ms (horizontal) and 50 mV (vertical, for action potential).
A and B: an atypical preparation in which retrograde AVN conduction was influenced to a greater degree by postganglionic vagal stimulation (PGVS) than antegrade atrioventricular nodal (AVN) conduction. Organization of records was as follows (from top to bottom): crista terminalis (CrT) electrogram, lower interatrial septum (IAS) electrogram, action potentials from AN region, and His bundle electrogram (H). Snort bar above H electrogram represents PGVS (APh 250 ms, duration 100 ms, amplitude 50 μA) and numbers below the H electrogram indicate conduction time (in ms). Preparation was driven at a cycle length of 450 ms. A shows antegrade conduction and I retrograde conduction. Note that fragmentation of action potential was induced by PGVS during retrograde conduction (B) which could account for longer retrograde that antegrade conduction time. C and D: same preparation and organization of records as A and B. In addition, a simultaneous recording of action potential from an N cell was added and amplitude of PGVS was incresed to 100 μA. Note that PGVS-induced hyperpolarization in N cell was greater during antegrade conduction (C). In contrast, the AVN conduction (D), and AVN block was observed distal to the impaled AN cell during the second cycle after PGVS. *Spontaneous sinus beat. See text for details. Calibration: 100 ms (horizontal) and 50 mV (vertical, for action potential).
The major mechanistic role of the orientation of the conducted wavefront in the differential effects of PGVS is further demonstrated in Fig. 8. In this case, a long train of antegrade (Fig. 8, A and B) or retrograde driving (Fig. 8, E and F) stimuli was interrupted by a stimulus with equal coupling interval but from the opposite site: retrograde in Fig. 8, C and D, and antegrade in Fig. 8, G and H. PGVS was introduced in Fig. 8, B, D, F, and H, with the same amplitude and optimal absolute phase. The latter was chosen to demonstrate the maximal effects at this amplitude of PGVS.
It is clear from the comparison of Fig. 8, B and D, that PGVS-induced hyperpolarization in the AVN should have been identical up to the introduction of the last beat (the same conclusion is valid for the situation presented in Fig. 8, F and H). Thus the differences in the PGVS-induced prolongation of the conduction time of the last beat should reflect primarily the effect from the changed orientation of conduction. As shown in Fig. 8, the first retrograde beat was accompanied by diminished effect of PGVS on the prolongation of the conduction time (100 to 117 ms, Fig. 8, C and D) as compared with the same effect during steady-state antegrade conduction (83 to 114 ms, Fig. 8, A and B). During subsequent steady-state retrograde conduction, PGVS effect further diminished (115 to 122 ms, Fig. 8, E and f), which was as expected from the lesser hyperpolarization (shown in Fig. 5). However, in this circumstance the first antegrade beat was accompanied by an increase in magnitude of the PGVS effect (90 to 115 ms, Fig. 8, G and H), thus demonstrating the same relationship independently of the order in which the switch in the orientation of the conducted wavefront was produced. Similar observations were found in four preparations in which the above type of perturbations were explored.
DISCUSSION
A major observation in this study was the demonstration that similar postganglionic vagal stimulation produced a more marked hyperpolarization in the AVN cells of the rabbit heart tissue preparation during antegrade than during retrograde conduction. Generally, this was accompanied with an overall greater effect of PGVS on antegrade vs. retrograde AVN conduction time.
Evidence for differences in the organization of AVN conduction based on the direction of propagation has been reported both in animal models 6) 7) 8) 26) 27) and in humans 2) 19). It is generally accepted that conduction delay in the AVN is greater during retrograde rather than antegrade propagation.
Advantages and Limitations of Methods Used in Present Studies
The data presented in this study show that, at comparable cycle lengths, the retrograde AV nodal conduction time in the rabbit heart preparation was usually longer than the antegrade; the reverse relationship was not observed in our experiments (Fig. 4). It is important to mention that in our model the atrioventricular nodal conduction time was measured as the interval between the earliest "input" (crista terminalis for the antegrade and His bundle for the retrograde mode, respectively) and the earliest "output" (the His bundle for the antegrade and crista terminalis for the retrograde mode, respectively), which are the areas closest to the AVN.
This method of measurement assured a more accurate estimation of the true nodal delay in comparison to the studies in patients or animals in which the atrioventricular interval was measured between the inscriptions of a surface atrial electrogram and the QRS complex 7), between a high right atrial electrogram and the His bundle electrogram 13), or between the right atrial appendage electrogram and the right ventricular electrogram 25) 28). Furthermore, in vivo, ventricular pacing itself could alter hemodynamic parameters and consequently the functional properties of VA conduction 5) 22) 23). Therefore, in such animal and human studies in vivo there are potential methodological limitations to precisely elucidating the AVN conduction time in both directions as well as to identifying the influence of vagal effects on these conduction properties 2).
FIG. 8.
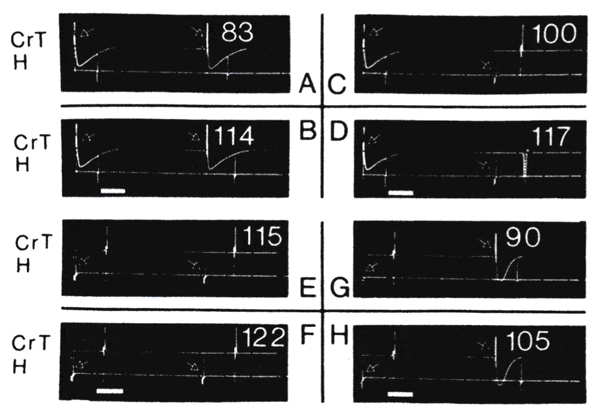 Differences in effects of similar bursts of postganglionic vagal stimulation (PGVS) introduced at steady state with an antegrade drive cycle length of 500 ms (A and B) and just before introduction of a retrograde stimulus with same coping interval (C and D). In both cases PGVS with duration 100 ms (short horizontal bar) was introduced at optimal phase, i.e., producing maximal effects. Arrows indicate drive stimuli. Numbers indicate corresponding conduction times in ms. Note that PGVS-induced prolongation of antegrade conduction time (A and B, 83 to 114 ms) was attenuated during retrograde beat (C and D, 100 to 117 ms). There is a similar organization of records and same parameters of PGVS as in previous panels, but steady-state retrograde train (E and F) was followed by an antegrade beat (G and H). Note that although PGVS effect was diminished during retrograde train (115 to 122 ms in E and F compared with 100 to 117 ms in C and D), this effect was again augmented with first antegrade beat (90 to 105 ms in G and H). See text for details.
Differences in effects of similar bursts of postganglionic vagal stimulation (PGVS) introduced at steady state with an antegrade drive cycle length of 500 ms (A and B) and just before introduction of a retrograde stimulus with same coping interval (C and D). In both cases PGVS with duration 100 ms (short horizontal bar) was introduced at optimal phase, i.e., producing maximal effects. Arrows indicate drive stimuli. Numbers indicate corresponding conduction times in ms. Note that PGVS-induced prolongation of antegrade conduction time (A and B, 83 to 114 ms) was attenuated during retrograde beat (C and D, 100 to 117 ms). There is a similar organization of records and same parameters of PGVS as in previous panels, but steady-state retrograde train (E and F) was followed by an antegrade beat (G and H). Note that although PGVS effect was diminished during retrograde train (115 to 122 ms in E and F compared with 100 to 117 ms in C and D), this effect was again augmented with first antegrade beat (90 to 105 ms in G and H). See text for details.
On the other hand, vagal effects on AVN conduction were evaluated in the present study by using short bursts of locally applied PGVS rather than by longer-lasting stimulation of the entire vagal trunk. This allowed evaluation of vagal effects in different regions of the AVN during both antegrade and retrograde conduction. Moreover, by using short bursts of PGVS (rather than a prolonged PGVS, which produced marked AVN block as in Ref. 29)), it was possible to study the phasic vagal effects and compare them in the antegrade vs. retrograde mode. This approach has a physiological basis, since it is accepted that phasic vagal control of the heart 12) is an important mechanism closely related to the bursttype efferent vagal discharges in response to alterations such as the rhythmic changes in the systolic blood pressure 9) 10).
Nevertheless, the results reported here should be analyzed within the limits of the methods used. Specifically, a comparison with the results obtained in clinical studies or with different cholinergic interventions (e.g., superfusion with acetylcholine) should be made with caution.
Antegrade vs. Retrograde AVN Conduction at Low Level of Vagal Activity
The nature of the directional difference in AVN conduction is not fully understood. However, based on the inhomogeneous morphology of the node 11) 24) and the inhomogeneity of its electrophysiological properties, one can speculate that electrotonic influences between the elements in the conduction pathway might be strongly dependent on the direction of conduction. It has been established 20) 27)that the configuration of action potentials of AVN cells may be different during antegrade and retrograde conduction (see also Fig. 7). The “convergent-divergent" geometry model used by Van Capelle and Janse 26) showed a good approximation with AVN behavior in vitro and was indirectly supported by the observations of Merideth et al. 18), which demonstrated that cellular excitation in the AVN was critically dependent on the characteristics of the wavefronts and their summation 17) 30). Thus successful depolarization of AVN cells, especially in the N region of the node, appears to depend at least on 1) inherent membrane properties, 2) characteristics of the current source, and 3) electrotonic communications with neighboring cells. The last two factors may differ depending on the orientation of propagation of the wavefront through the node.
The resting autonomic vagal tone, by its depressant effect, might modify antegrade vs. retrograde properties of the AVN. However, there is evidence that local atropinization of the AVN region did not restore blocked retrograde conduction 7). Human studies 4) using the transplanted heart showed results similar to those in the innervated heart, suggesting that the autonomic nervous system appears to have little influence on the resting basic asymmetrical properties of the atrioventricular conduction system. In addition, even when atropine restored retrograde conduction in some patients with unidirectional retrograde block 3), a profound difference between antegrade and retrograde conduction times still remained. The data from our study, in which the level of background autonomic vagal tone should be considered negligible 29), also supports the view that the asymmetric properties of AVN conduction leading to "better" antegrade and "worse" retrograde conduction cannot be explained by a different, i.e., directionally dependent influence of the resting vagal tone.
Antegrade vs. Retrograde AVN Conduction During Increased Vagal Activity
The data from the present study indicate further, that an increase in vagal tone (e.g., as simulated by PGVS) can influence antegrade vs. retrograde AVN conduction differently.
It appears that the change in direction of conduction leading to entirely different intranodal organization of excitation (i.e., electrotonic environment) is a major factor underlying the differential effects of PGVS. In fact, the N region, similarly depressed by PGVS, can be transversed differently antegradely vs. retrogradely depending on the effective strength of the current source in both directions, as suggested from the results in Fig. 8. Additionally, the change in the direction of conduction was accompanied by modulation of PGVS-induced hyperpolarization. One can assume that the amount of the released mediator, acetylcholine, and its subsequent hydrolysis, depend on the intensity of PGVS but not on the direction of propagation. However, the degree of PGVS-induced hyperpolarization should depend not only on the amount of the mediator in contact with the cellular membrane, but also on electrotonic factors reflecting the activation status of neighboring cells and their level of depression by PGVS. Thus during antegrade conduction the process of PGVS-induced hyperpolarization of a given N cell will reflect, in part, the 'convergence" of the propagated wavefront toward the NH region. In contrast, during retrograde conduction, hyperpolarization of the same cell will reflect, in part, "divergence" of the propagated wavefront as it moves toward the AN region. However, the particular quantitative differences are difficult to explain. A detailed knowledge of intercellular organization of this part of the node is necessary to speculate about the reasons leading to a greater PGVS-induced hyperpolarization of the N cells during antegrade rather than retrograde conduction.
Based on the observations in the present study, a role for increased vagal tone in the occurrence of unidirectional retrograde AVN block in vivo cannot be excluded. In fact, as previously shown, vagal influence in the AVN can produce increased inhomogeneity 15), which will influence the properties of conduction in addition to, or in conjunction with, the changes caused by hyperpolarization itself. Depending on the intensity, timing, and location of PGVS, the level of disorganization produced in the AVN may differ, also being dependent on the direction of propagation of the wavefront, as shown in Fig. 7.
However, the development of AVN block does not reliably represent the directional-dependent differences in AVN conduction resulting from increased vagal tone. In fact, it is obvious that AVN block occurs after the conduction delay has reached some critical maximal limit. The latter is apparently functional and dependent on the direction of propagation among other factors. Whether block is observed first (i.e., at lower PGVS amplitude) in the antegrade or retrograde direction depends on the basic asymmetric properties of AVN conduction in certain species and not merely on the absolute effect of vagal stimulation (hyperpolarization, absolute increase in the conduction delay). For example, if the maximal achievable conduction time in the retrograde mode in the case illustrated in Fig. 3 were 120 ms, then block would have occurred first in retrograde conduction, although the conclusion that the antegrade conduction was relatively more influenced by PGVS would still be correct. Importantly, unidirectional retrograde block before PGVS was not observed in any of the rabbit heart proparations reported in the present study, whereas unidirectional retrograde block has been frequently observed in the dog 7) and humans 1) 13). Thus the results obtained during analysis of vagal-induced retrograde block 7) 13) 29) should not be regarded as contradictory to the data presented in this study.
On the other hand, the data of Urthaler et al. 25) obtained with continuous vagal stimulation in dogs showed that by increasing the frequency of stimulation similar depression of both antegrade and retrograde AVN conduction was observed. In the dog model with phasic vagal stimulation, Wallick et al. 28) demonstrated that the direction of conduction did not affect the amplitude of the phasic response curve, although the maximal conduction time was longer during retrograde direction. We assume that the difference between the results presented in the present study and those in previous studies 25) 28) might represent certain species variations in the basic asymmetric properties of AVN conduction.
Our study suggests that the observed differential effects of PGVS on antegrade vs. retrograde conduction reflect predominantly the influence of the orientation and homogeneity of the wavefront conducted through the AVN. These effects are accompanied by directionally dependent modulation of PGVS-induced cellular hyperpolarization. The precise mechanisms underlying these interrelationships deserve further investigation.
We gratefully acknowledge the technical help of Carl Hurt and Rohn Price and the assistance of Rose Marie Wells and Marilyn Lucianetti in preparation of this manuscript.
E. L. Mlcheleon was supported in part by Clinical Investigator Award K08 HL-01312 from the National Heart, Lung, and Blood Institute.
Address for correspondence: T. Mazgalev, Cardiovascular Division, The Lankenau Medical Research Center, Lancaster, west of City Line Ave., PhiladelPhia, PA 19151.
Received 22 December 1986; accepted in final form 10 June 1987.
REFERENCES
- Akhtar, M. Retrograde conduction in man. PACE 4:548-562, 1981.
- Akhtar, M., A. N. Damato, W. P. Batsford, J. N. Ruskin, and J. B. Ogilkelu. A comparative analysis of antegrade and retrograde conduction patterns in man. Circulation 52:766-778, 1975.
- Akhtar, M., M. Shenasa, S. Denker, S. Patel and J. Harris. Reversible nature of unidirectional (retrograde) atrioventricular nodal block in the human heart (Abstract). Clin. Res. 30: 758A, 1982.
- Bexton, R. S., A. W. Nathan, K. J. Hellestrand, R. Corypearce, R. A. J. Spurrell, T. A. H. English, and A. J. Camm. The electrophysiologic characteristics of the transplanted human heart. Am. Heart J. 107: 1-7, 1884.
- Blanc, J. J., E. Gestin, D. Guillerm, J. Boschat, and P. Penther. Response of normal and abnormal sinus node to right ventricular stimulation. Am. J. Cardiol. 48: 429-436, 1981.
- Cranefield, P. F., B. F. Hoffman, and A. Paens De Carvalho. Effects of acetylcholine on single fibers of the atrioventricular node. Circ. Res.7: 19-23, 1959.
- Fransis, C. K. and H. E. Hoff. The influence of local atropinization of the A-V node on retrograde V-A conduction . cardiovasc. Res. Cent. Bull 5-13, 1965.
- Hoffman B. F., and P. F. Cranefield. Electrophysiology of the Heart. New York: McGraw-HIll, 1960, p.150.
- Iriuchijima, J., and M. Kumada. Efferent cardial vagal discharge of the dog in response to electrical stimulation of sensory nerves. Jpn. J. Physiol. 13: 599-605, 1963.
- Iriuchijima, J., and M. Kumada. Activity of single vagal fibers efferent to the heart. Jpn. J. Physiol. 14: 479-487, 1964.
- Janse, M. J., F. J. L. Van Capelle, R. H. Anderson, P. Touboul, and J. Billette. Electrophysiology and structure of the atrioventricular node of the isolated rabbit heart. In: The Conduction System of the Heart, edited hy H. J. J. Wellens, K. I. Lie, and M. J. Janse. The Hague: Nijhoff, 1978, p.296-315.
- Levy M. N., and P. J. Martin. Neural control of the heart. In: Handbook of Physiology. The Cardinvascular Systern. Thc Heart. Bethesda, MD: Am. Physiol. Soc., l979, sect. 2, vol. I, chapt. 16, p.581-620.
- Mahmud, R., S. T. Denker, M. H. Lehmann, A Addas, and M. Akhtar. Unidirectional retrograde atrioventricular nodal block in man: determinants of reversibitity by vagal antagonism. Am. Heart J. 110: 568-574, 1985.
- Mazgalev, T., L. S. Dreifus, E. S. Michelson, and A. Pelleg. Vagally induced hyperpolarization in the atrioventricular node. Arn. J. Physiol. 251 (Heart Circ. Physiol, 20): H631-H643, 1986.
- Mazgalev, T., L. S. Dreifus, E. S. Michelson, and A. Pelleg. Effects of postganglionic vagal stimulation on the organization of atrioventricular nodal conduction in isolated rabbit heart tissue. Circulation 74: 869-880, 1986.
- Mazgalev, T., L. S. Dreifus, E. S. Michelson, A. Pelleg, and R. Price. Phasic effects of postganglionic vagal stimulation on atrioventricular nodal conduction. Am. J. Physiol 251 (Heart Circ. Physiol. 20): H619-H630, 1986.
- Mendez, C. Characteristics of impulse propagation in the mammalian atrioventricular node. In: Normal and Abnormal Conduction in the Heart, edited by A. Paes de Carvalho, B. F. Hoffman, and M. Lieberman. Mt. Kisco, NY: Futura, 1982, p.363-377.
- Merideth, J., C. Mendez, W. J. Mueller, and G. K. Moe. Electrical excitability of atrioventricular nodal cells. Circ. Res. 23: 69-85, 1968.
- Narula, O. Retrograde pre-excitation. Comparison of antegrade and retrograde conduction intervals in man. Circulation 50: 1129- 1143, 1974.
- Paes De Carvalho, A. Excitation of atrioventricular node during normal rhythm. Effects of acetylcholine. In: Mechanisms and Therapy of Cardiac Arrhythmias. edited by L. S. Dreifus and T. H. Moyer. New York: Grune & Stratton, 1966, p.341-352.
- Pasquariello, J. L., R. J. Hariman, J. A. C. Gomez, P. Barreca, and N. El-Sherif. Autonomic dependence of ventriculo-atrial conduction (Abstract). J. Am. Coll. Cardiol. 3: 517, 1984.
- Paulay, K., and A. D. Damato. Comparison of atrial and ventricular drive on sinus nodal function in the dog. Am. J. Cardiol. 31:41-46, 1973.
- Schuilenburg, R. M. Patterns of V-A conduction in the human heart in the presence of normal and abnormal A-V conduction. In: The Conduction System of tlle Heart, edited by H. J. J. Wellens, K. I. Lie, and M. J. Janse. The Hague: Nijhoff, 1978, p.485-503.
- Spach, M. S. Impulse propagation in the AV node. In: Normal and Abnormal Conduction in the Heart, edited by A. Paes de Carvalho, B. F. Hoffman, and M. Lieberman. Mt. Kisco, NY: Futura, 1982, p.377.
- Urthaler, F., B. N. Neely, G. R. Hageman, and L. R. Smith. Differential sympathetic-parasympathetic interactions in sinus node and AV junction. Am. J. Physiol. 250 (Heart Circ. Physiol. 19): H43-H51, 1986.
- Van Capelle, F. J. L., and M. J. Janse. Influence of geometry on the shape of the propagated action potential. In The Conduttion System of the Heart, edited by H. J. J. Wellens, K. I. Lie, and M. J. Janse. The Hague: Nijhoff, 1978, p.316-335.
- Van Capelle, F. J. L., M. J. Janse, P. J. Varghese, G. E. Freud, C. Master, and D. Durrer. Spread of excitation in the atrioventricular node of isolated rebbit hearts studied by multiple microelectrode recording. Circ. Res. 3l: 602-616, 1972.
- Wallick, D. W., S. L. Stuesse, and Y. Masuda. Sympathetic and periodic vagal influence on antegrade and retrograde conduction through the canine atrioventriculer node. Circulation 73:830-836, 1986.
- West T. C., and N. Toda. Response of the AV node of the rabbit to stimulation of intracardiac cholinergic nerves. Circ. Res. 20: 18- 31, 1967.
- Zipes, D. P., C. Mendez, and G. K. Moe. Evidence for summation and voltage dependency in rabbit atrioventricular nodal fibers. Circ. Res. 32: 170-177, 1973.


